本論では、これまで一貫して「筋肉は熱揺らぎを利用している」ことを主張してきたが、 もう少し公平な立場から見て、現在のところ筋肉のメカニズムがどのように 理解されているかについてまとめておこうと思う。 ここでは 「分子細胞生物学(下) 第4版 H.Lodish他, 東京科学同人(2001)」 という本を参照した。 以下に示す図は、同書の 「18・3 ミオシン:アクチンモータータンパク質」 にあったものを筆者が書き写したものだ。(直接コピーするのは版権があるので)
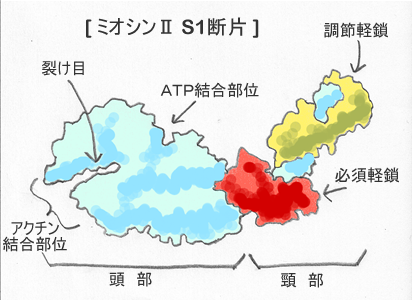
1つ目の図は、ミオシン頭部の模式図である。 筋肉のミオシンⅡには、この頭部が2つある。 頭部には裂け目があり、その先端がアクチンと結合する場所になる。 裂け目の反対側にATPと結合する場所がある。 2つの結合部位の距離は 3.5nm である。
以下は同書からの引用。
「アクチンやATPと結合したり離れたりすることによって
頭部の裂け目が開いたり閉じたりして、
頸部を支点として頭部に回転を起こさせるのではないだろうか。」
「ここで述べるモデルでは、ATP1分子の加水分解とアクチンフィラメントに沿って
ミオシン分子が一歩動くことが共役しているという仮定がある。」
「しかし、加水分解と運動が密接に共役している証拠は決定的となっていない。
ATP1分子の加水分解ごとにミオシンが1歩ずつ動くこともあり、
それ以上動くこともあるようである。」
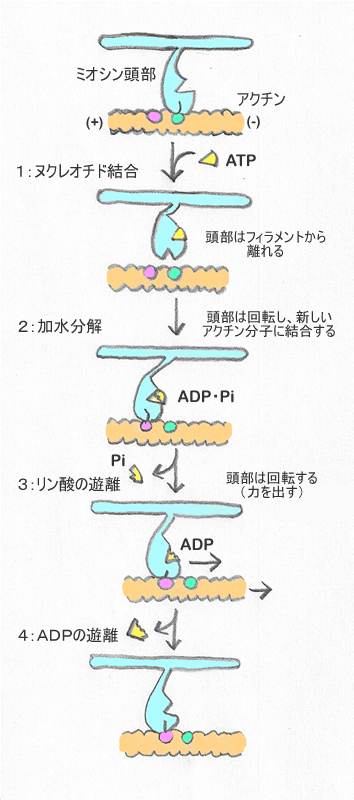
2つ目の図は、1つ目の図にあるミオシン頭部の構造をもとに考えられた、 筋肉のすべり運動のメカニズムである。 ATPがADP+リン酸に加水分解するエネルギーを使って、 アクチン繊維を図の右方向に動かす様子が描かれている。 加水分解のエネルギーは、主として「頭部を回転する(力を出す)」過程に使われている。
タイトカップリング対ルースカップリングといった構図から言えば、 ここでの説明はタイトカップリング派である。 そして、こうした立派な教科書に載っていることからすると、 どちらかといえば、この図に描かれているものが「世界標準的な」理解なのだろう。 ちなみに、この教科書には熱揺らぎとか、ブラウン運動の利用といった記載は (筋肉の運動に関しては)全くない。
それでは、ルースカップリングの立場から筋肉の運動を説明すると、 どのようになるだろうか。 ミオシン頭部の構造が1つ目の図のもので間違いないとして、 もしミオシンが熱揺らぎを利用していたとすると、 具体的にどの過程で熱揺らぎが関与しているのだろうか。 熱揺らぎによってアクチンが運動し得るのは、 ミオシンとアクチンが離れているときである。 図で言えば1:と2:の間に相当する。 ここで熱ゆらぎが働けば、ミオシンがアクチンの上を、 場合によっては数ステップ、行ったり来たりすることになるだろう。 ミオシンの運動を調べると、実際そのようになっているらしい。
しかし、熱揺らぎが左右均等に働いたならば、アクチンが右に動くか左に動くか分からないので、 結果として熱揺らぎは筋肉の運動全体に全く寄与しないことになる。 そこでミオシンはATPを使って、 左なら左といった具合に一方向に動く確率を高めているのだろう、 という推測が成り立つ。 いわゆるバイアスド・ブラウン運動モデルは、こうした推測から考えられたものだ。
さて、ここから先のお話は、何か実験的な根拠に基づいたものではなく、全て推測となる。 以下は素人の推測の域を出ないお話として受け止めてほしい。
もしミオシンが不確定分子モーターの具現化と言うのであれば、
それは具体的にどういった構造をとっているのだろうか。
筋肉のすべり運動を表す2つ目の図は、
「ATPが結合した結果 => ミオシンが動く」
ものとして描かれている。
ここで1つ発想を逆転させて、この原因、結果の矢印 => の向きを
反対にしてみたらどうなるだろう。
つまり、
「たまたまミオシンが動きそうになったとき => ATPが結合する」
と捉えるのである。
「たまたまミオシンが動きそうになる」とはどういうことか。
それは、熱揺らぎによってミオシンが図の左向きにゆさぶられたときである。
ミオシンに左向きの力が加わったとき、ATPはミオシンに結合し易くなる。
すると、アクチン:ミオシン間の結合が切れて、その後しばらくの間
アクチンは右向きに、ミオシンは左向きに運動する。
加水分解が起こって、再びアクチン:ミオシンが結合したときには、
結果としてアクチンは大きく右に動いたことになる。
このメカニズムであれば、アクチンはミオシンの長さ1個分ではなく、
それよりもずっと長距離動くことができるだろう。
つまり、ミオシンは自らの腕力に物を言わせて力ずくでアクチンを動かすのではなく、 相手の出方を見て、それを上手に利用しているのである。 そしてこの場合ATPの結合は、相手の出方を探る観測装置として働いている。 たまたま左向きにゆさぶられたときATPが結合しやすくなる、 というメカニズムが、結果として熱揺らぎ運動の観測装置として機能する。 加水分解の自由エネルギーは、観測装置が逆向きに動作しないことを保証する。 全プロセスを通じて逆向きに動作しなければよいのだから、 必ずしも加水分解のタイミングに合わせて出力が取り出されなくてもよい。 また、エネルギーを観測装置だけに振り向ければよいのだから、 力ずくで直接アクチンを動かすよりも、わずかの力で済むことになる。 ATP自身の持つエネルギーに加えて、熱エネルギーを取り込むことが可能となるのだ。
相手の出方に合わせて力を利用する。 日本人の私としては、ここで「柔道」を想起せずにはいられない。 柔よく剛を制す。 マックスウェルの悪魔は柔の道を究めていたのである。
もし以上の推測が正しければ、筋肉は間違いなく不確定分子モーターの実例であったことになる。 なぜなら、たまたま熱揺らぎが目的の方向に働いたときに運動するのであれば、 その出力タイミングは不確定となるはずだからである。 そして「出力が不確定な分だけ熱揺らぎを利用している」ということは、 不確定分子モーターの主張と一致する。
さて、ここまでの推測の上にさらなる推測を重ねて、次に筋肉全体のことを考えてみよう。 筋肉は、非常に数多くのミオシン:アクチン分子モーターから成り立っている。 このことは、熱揺らぎの利用という観点からすると、また1つの難しい問題をはらんでいる。 ここで運動会の綱引きを想像してみてほしい。 綱引きというものは全員が一致団結して、一斉に同じタイミングで引っ張った方が力強い。 各自がてんでばらばらに引っ張っても、いまひとつ有効に働かないのである。 もし個々の分子モーターが熱揺らぎによって不確定に動いていたならば、 それは綱引きと同じで、全体として効率よく機能しないことになる。 せっかく分子サイズで熱揺らぎを利用していたとしても、 それらが多数集まったときに、熱揺らぎを利用するメリットが相殺されてしまうのである。
そこで考え直したいのは、 「筋肉というものは、単なる分子モーターの寄せ集めなのだろうか」 ということである。 綱引きの例で言えば、みんなが気持ちを1つにすることによって、いつのまにか自律的なリズムが生まれる。 そうしたリズムが生まれると、全体として非常に強力なパワーを発揮する。 これと同じように、筋肉分子モーターも単純な寄せ集めではなく、 全体でシンクロするような仕組みがあるのではないかと思う。 1個の分子モーターが熱揺らぎを利用している、というだけでも充分驚きだが、 それらが集まって互いにシンクロしている、というのはさらなる驚きであろう。 しかし、生物はそれをやってのけている気がする。
柔道に綱引き。 生体の中は、思った以上ににぎやかな所のようだ。



